THE USE OF GENETICS IN BREEDING CHAMPIONS
PART 6: CHROMOSOMES AND THEIR BEHAVIOUR CONTINUED
INTRODUCTION
In part 5 we discussed most of the “interactions” and how we can find out if certain genes, or at least traits, are present in our pigeons. We need to know this before we can decide which hens and cocks should be mated with one another.
In this part we will start relating genes and traits to characteristics as we see them in our pigeons.
THE RELATIONSHIP OF GENES TO TRAITS
No gene affects the phenotype in an instantaneous fashion. Rather, a gene’s effects are an aspect of the phenotype that is influenced by metabolic pathways in which the product of each particular allele is active.
A single protein may be active in several metabolic pathways in the body. A change in the gene influencing the production of a protein will therefore affect all pathways in which that protein operates. The net result may be visible as a change in the phenotype. However, many mutations are not detected because they have only a minor effect on the protein structure. Therefore, knowledge of changes in the genotype depends on the observer’s ability to detect their effect on the phenotype. When an observable change occurs, it may be seen in a single trait or in several, that is, as a pleiotropic effect.
We already said that one should be careful of judging the racing and breeding value of a pigeon based on visible (phenotypical) traits. Then again, we don’t have much else to go on (except for racing results, which should always be the most important yardstick). Experience showed that there are some visible traits that are mostly found in top class racers. The importance of each trait is dependent on the racing distance at which we would like our pigeons to perform best. Pigeon fanciers will always argue about the merits of classifying pigeons as short, middle or long distance racers. Fact remains, some pigeons do tend to perform better at particular distances. In fact, the observant fancier will notice that most of his racers also perform better under particular environmental conditions, time of the day, time of the year, etc.) You do not need to be a scientist or have sophisticates equipment and laboratories at your disposal to identify the following traits:
- Speed.
- Absence of physical flaws.
- Strong frame (bone structure).
- Supple muscles.
- Strong character – this can be gauged from the pigeon’s fighting spirit, will-power, urgency to trap, etc.
- Intelligence – this can be seen in the pigeon’s eyes and behaviour.
- Good oxygen uptake – this can to some extent be derived from the size of the pigeon’s chest.
- Balanced wing with a short forearm.
It is not only the strong traits that you must seek in your pigeons – the weak traits also need to be identified (and rooted out). You should make notes of negative observations, e.g. a good racer that constantly performs poorly in bad weather conditions or a breeder that fails to transfer good traits. External factors should also be taken into consideration, e.g:
- The fancier (if you plan on obtaining pigeons from someone else) – does he have the ability to perform well with average pigeons, is he patient, is he honest, etc?
- The loft – loft position relative to other lofts in the club, federation or union.
- Training method.
- Feeding method and feeding mixtures.
With the exception of a few DNA tests, which we as pigeon fanciers cannot use anyway, we cannot know the genetic makeup of our pigeons, only the physical makeup, or phenotype. We tend to break that phenotype up into traits. Human beings define all of these “traits”. Very few of them actually refer to single genes that might be inherited as dominant, recessive, incomplete dominant or co-dominant. In general we use dominant, recessive, co-dominant or intermediate (incomplete dominant) to refer to genes at the same location on a single pair of chromosomes, i.e., alleles at the same locus. There are cases where genes at one locus can “hide” genes at another locus. An example is recessive white eye-colour being hidden by dominant yellow eye colour. This type of relationship among different loci is called epistasis. The locus of the hidden gene is referred to as hypostatic. In some cases an epistatic locus has an allele that allows the hypostatic locus to show its effects.
Sometimes we break down a trait into a specific combination of genes. In the case of colour, for instance, we know of a considerable number of genes that affect colour through specific processes. In some cases, this knowledge has fed back on what we consider to be traits. In addition, a pigeon’s colour can be affected by minor genes, (such as the modifier genes determining the shade of a particular colour) by random factors (which probably influence the exact pattern and location of colour spots) and by environmental factors (such as the loft layout, nutrition, exposure to the sun, etc). The point is that very few of the traits that humans have chosen are in fact due solely to the effect of a single pair of alleles at a single locus.
The size of the pigeon, form of the wing, shape of the muscles are probably not single-gene traits, but rely on the interaction of several pairs of genes, with perhaps some influence from the environment. (The same probably apply to traits that we cannot physically see, such as perseverance, speed, etc.) We will return to this when we discuss polygenic traits.
Dihybrid crosses. A dihybrid cross involves a study of inheritance patterns for organisms differing in two traits. A dihybrid cross starts with two parents who are pure breeding for alternative forms of two traits. As we saw in the second and third articles, Mendel invented the dihybrid cross to determine if different traits of pea plants, such as flower colour and seed shape, were inherited independently. (Http://www.biology.arizona.edu/mendelian_genetics/problem_sets/dihybrid_cross.) The complications of codominance, lethality, variable expressivity, and incomplete penetrance also may effect the outcome of dihybrid crosses. If one trait has a lethal associated with it, then any phenotype having the lethal genotypic combination would disappear from the F2 generation, thus changing the ratio observed.
Types of inheritance. We will consider a number of types of inheritance in a later article on genetics. The first group actually refer to single-gene traits. Any of these types of inheritance may also be involved in the inheritance of multi-gene traits.
Allele gene pairs. This is a very important concept. In cells we find pairs of inherited traits – one from the father and the other from the mother. This is called an allele gene pair. This is, furthermore, the concept with which we can differentiate between homozygosity and heterozygosity in simple terms. Suppose both the cock and the hen carry the gene for the colour blue in their chromosomes. Seeing that colour is linked to sex, both the male and female offspring will be blue. This is an allele gene pair, and we call it a homozygote allele pair.
Should one parent carry the gene for red and the other for blue, the allele gene pair in the offspring will consist of two different traits for colour. We call this a heterozygote allele pair.
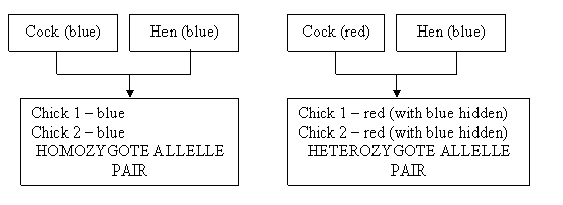
Figure 9: Homozygote vs heterozygote allele pairs.
So, allele gene pairs are gene-qualities inherited from both parents, which can be either homozygote or heterozygote.
Dr Jaap Nel. Email: jaap7@iafrica.com.